- Nature Chemical Biology, 8, 966–968 (2012) doi:10.1038/nchembio.1109
☆ O-GlcNAc転移はどう起こる? その2
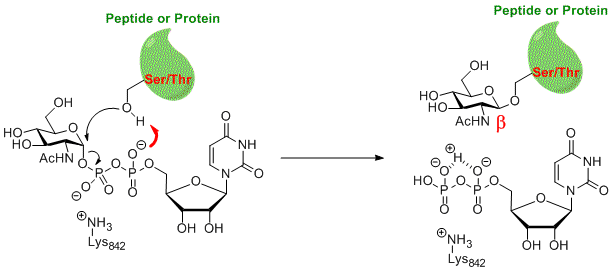
彼らは触媒反応の最初と最後の結晶構造を比較することにより触媒機能を明らかにしようと考えましたが、その鍵となったのはUDP-5SGlcNAcでした。通常のUDP-GlcNAcでは結晶化の間に転移反応が進行してしまい、反応の初期段階の結晶構造が得られなかったのです。このUDP-5SGlcNAcはOGTの阻害剤として既に知られており、天然型UDP-GlcNAcと比べ圧倒的に基質になりにくい性質を利用しています。

○求電子的転移機構で反応が進行する(糖加水分解酵素でよく見られる反応機構)
○水分子を介してAsp554が水酸基の水素を引き抜く
他のポイントとしては
○Lys842,Thr921,糖アセトアミドが安定化に寄与
○His498,558は距離が遠すぎて引き抜きは不可能、Tyr841も同様。
○alpha-リン酸基は3.5A以内にあり、距離的には引き抜き可能に見えるが、pKaを考えると難しい。また、その場合はアノマー位への攻撃が難しい位置関係になってしまう。
両論文とも得られた構造は非常に似ているように感じましたが、結晶構造の解釈の違いで結論が違うものになっています。現時点でどちらが正しいのか筆者にはなかなか判断がつきませんが、ひとまず後続の論文を楽しみに待ちたいと思います。